
Патрон: Александр Демидов
Temet Nosce
В поисках ответов на свои вопросы главный герой трилогии сестёр Вачовски, Нео, приходит к специалисту – пифии. В кадр попадает табличка над ее дверью с надписью Temet nosce, лучшим в мире слоганом для уважающего себя оракула. Если верить греческому географу и путешественнику Павсанию, то это одна из трёх максим, которые были начертаны на входе в храм Аполлона в Дельфах. Temet nosce или nosce te ipsum, это латинский вариант греческого gnōthi seautón (γνῶθι σεαυτόν), “познай себя” (know thyself).

Глубина и многозначительность фразы позволяет множество трактовок. В “Прометее прикованном” Эсхила она звучит как поучение мятежному полубогу, что хорошо бы знать своё место и не зарываться. Платон в своих диалогах неоднократно использует её, приписывая Сократу в самых разных контекстах. “Познай себя” мелькает и в “Гамлете” Шекспира, и в “Левиафане» Гоббса, и в линнеевской “Systema Naturae”, и в “Альманахе бедного Ричарда” Бенджамина Франклина, и ещё много где. Что до ответа на вопрос, зачем, собственно, познавать себя, можно принять два варианта, на ваш выбор.
Сократ. Посмотри и ты: предположим, что изречение это советовало бы нашему глазу, как человеку: “Увидь самого себя!” Как восприняли бы мы подобный совет? Не так ли, что глаз должен смотреть на то, что позволит ему увидеть самого себя?Алкивиад: Это понятно.
Платон “Диалоги: Алкивиад”
„Знай врага и знай себя: тогда в тысяче битв не потерпишь поражения.“
Сунь Цзы “Искусство войны”
Чеширский Кот на вопрос Алисы куда ей идти, ответил, что рано или поздно куда-нибудь попадёшь, если достаточно долго идти. Данте Алигьери, когда ему было тридцать пять лет тоже задавался вопросом, куда дальше – со строк “Земную жизнь пройдя до половины, я очутился в сумрачном лесу” начинается “Божественная комедия”. Чтобы познать, понять себя и “сориентироваться на местности”, может помочь взгляд в прошлое – кто мы и откуда пришли. Проследить историю человечества в глубь веков, на тысячи и миллионы лет назад, до момента когда наши предки откололись на эволюционном древе от шимпанзе, а оттуда уже вдоль стрелы времени – до появления Homo sapiens. Ответы на многие наши вопросы могут скрываться в толще нашего общего прошлого.
Филетический градуализм и прерывистое равновесие
Эволюцию как изменение во времени можно разделить на два элемента. Один элемент — это постепенные, постоянные изменения, от предка к потомку, от родителя к потомку, которые обеспечивают непрерывность жизни. Это классический, дарвиновский взгляд – непрерывный и кумулятивный процесс происхождения с модификациями. Благодаря непрерывным дебатам, чего в эволюции больше, и что главнее: градуализм или прерывистое равновесие, этот элемент принято считать нормальной эволюцией. Он настолько распространен и вездесущ, что не вызывает никаких сомнений в своей объективности. Она происходит постоянно, потому что вариации, мутации, изоляция, поток генов, дрейф (поток и дрейф генов – это разные вещи) и отбор неизбежно присутствуют.
Дрейф генов (genetic drift) При генетическом дрейфе частота аллелей (разных формы одного и того же гена, расположенных в одинаковых локусах парных хромосом) в популяции меняется в результате случайного отбора, когда не происходит адаптации. Генетический дрейф вызывают два механизма. Первый - это эффект бутылочного горлышка (bottleneck effect). Это генетический дрейф в популяции после того, как она пережила катастрофическое событие, например наводнение. Бутылочное горлышко происходит, когда частота аллелей основного признака в исходной популяции снижается, потому что погибло очень много особей, несущих этот аллель. В результате большая часть выжившей популяции вымирает, оставляя в живых несколько случайных особей. Другой механизм называется эффектом основателя (founder effect). Это когда несколько членов популяции отделяются и создают свою собственную группу. Из-за случайной выборки, создавшей новую группу, частота аллелей может резко измениться в зависимости от селективного давления, оказываемого на особей.
Поток генов (gene flow) отличается от генетического дрейфа тем, что представляет собой перенос аллелей или гамет из одной популяции в другую. Это происходит, когда популяция мигрирует или становится географически изолированной. Поток генов отличается от генетического дрейфа, наблюдаемого при эффекте основателя, когда новая группа формируется на территории, где нет существующей популяции.

Второй элемент — это более фундаментальная и радикальная сторона эволюционных изменений. Возникновение вида — это не просто еще одна мутация, это изменение ступени в эволюционирующей родословной. Есть разница между еще одним жуком и первым сухопутным позвоночным, или первым теплокровным существом. Однако, даже если смотреть шире, существует разница между эволюцией основных новых адаптаций и эволюцией совершенно новых биологических систем, таких как многоклеточность. Эти элементы можно назвать переходной эволюцией.
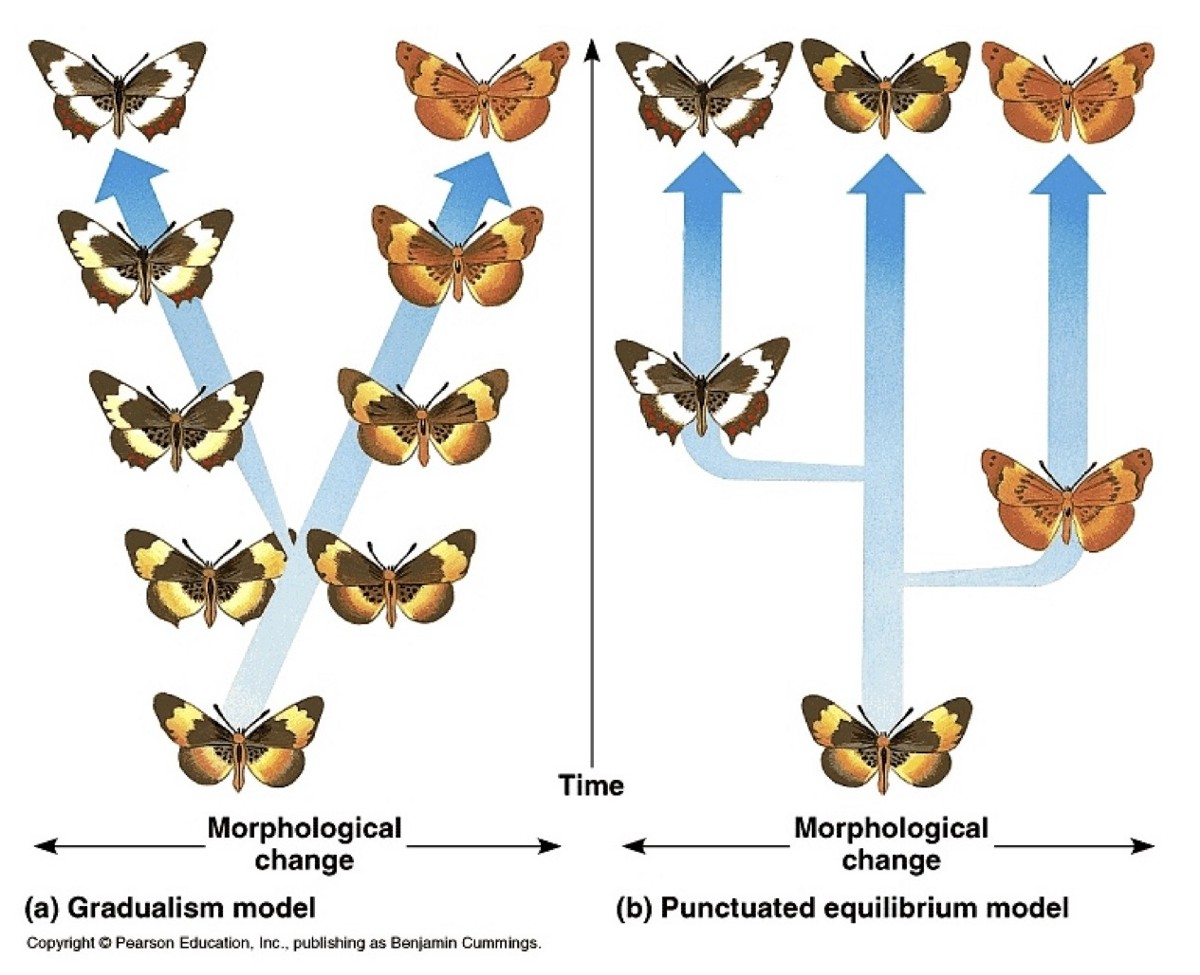
Дебаты и противоречия между сторонниками “нормальной и “переходной” эволюции ярче всего выражены в тянущемся уже которое десятилетие споре о “филетическом градуализме” и “прерывистом равновесии” – контрастирующих моделях эволюции, которые основываются на паттернах найденных ископаемых.
Прерывистое равновесие (punctuated equilibrium) – это морфологический стазис между резкими событиями видообразования. Все было тихо, спокойно и неизменно, пока резко не появились новые виды.
А филетический градуализм (phyletic gradualism) – это плавный (градуальный) процесс постепенных изменений в пределах рода (от греческого φυλή/φῦλον [phylé/phylon] «племя, клан, раса”), созвучный дарвиновскому взгляду, когда виды претерпевают более плавные непрерывные изменения.

Вера в то, что ископаемые виды оставались в морфологическом стазисе на протяжении всего своего существования, побудила Элдриджа и Гулда предложить эту модель в 1972 году в качестве альтернативы распространенной парадигме постепенной эволюции внутри родов (филетический градуализм). При филетическом градуализме эволюция происходит медленно и неуклонно, без особой ассоциации морфологических изменений с видообразованием; между периодами ветвления происходит столько же или даже больше эволюционных изменений, чем в самих точках ветвления. В отличие от этого, при прерывистом равновесии новый вид возникает быстро (прерывистость) и затем остается практически неизменным (равновесие или стазис) до конца своего существования.
Появление человека – это следствие постепенного накопления изменений, или результат взрывных, скачкообразных “прыжков” и ветвлений”? С одной стороны, нет сомнений в том, что люди представляют собой вид, значительно отличающийся от других приматов, и что их воздействие на биосферу было огромным и не только продолжает оставаться таковым, но и, вероятно, увеличивается. А с другой стороны, человеческая биологическая организация не так уж сильно отличается от организации шимпанзе и бонобо. Был ли в нашей эволюции тот “крупный переход”, который сделал нас теми, кто мы есть?
Четыре уровня эволюционных переходов
В эволюционном процессе можно выделить четыре фундаментальных типа изменений, которые могут быть иерархически вложены друг в друга. Это:
1. Базовая эволюция
2. Образование новых таксонов
3. Появление новых адаптивных зон
4. Глобальные эволюционные переходы

Базовая эволюция
Самый первый уровень – это возникновение новых черт благодаря мутациям, которое приводит к поэтапному изменению фенотипа. Вот она, эволюция в классическом, дарвиновском, понимании, когда накопление небольших изменений направляет траекторию эволюции и появление новых адаптаций. Появление новых точек или пятнышек, изменение расцветки покровов, которые происходят путем селекции или генетического дрифта. На этом этапе новые виды могут образовываться путём анагенеза – постепенного накопления небольших изменений в рамках скрещивающейся линии. Однако, анагенез случается куда реже кладогенеза.

Образование новых таксонов
На этом уровне происходит кладогенез (образование новых клад, от греческого κλάδος (kládos) ‘ветка’) – там, где раньше был один вид, появляется два, каждый из которых двигается дальше по своей эволюционной траектории. Кладогенез является фундаментальной основой биоразнообразия, и основным механизмом видообразования. Долгосрочная “фиксация” эффекта кладогенеза реализуется следующими механизмами:
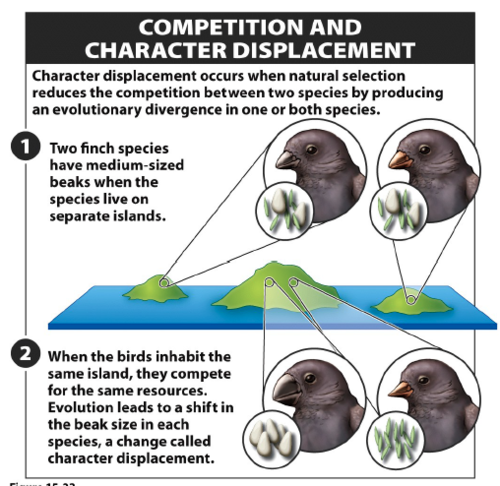
- Вытеснение признаков (character displacement) — изменения, которые происходят, когда два похожих вида обитают в одной среде. В таких условиях естественный отбор благоприятствует расхождению в характерах — морфологии, экологии, поведении и физиологии. Самый наглядный пример – знаменитые “дарвиновские” вьюрки обитающие на Галапагосских островах, у которых заметно различаются размеры тела и клюва, а также его форма, что подтолкнуло сэра Чарльза к размышлениям.
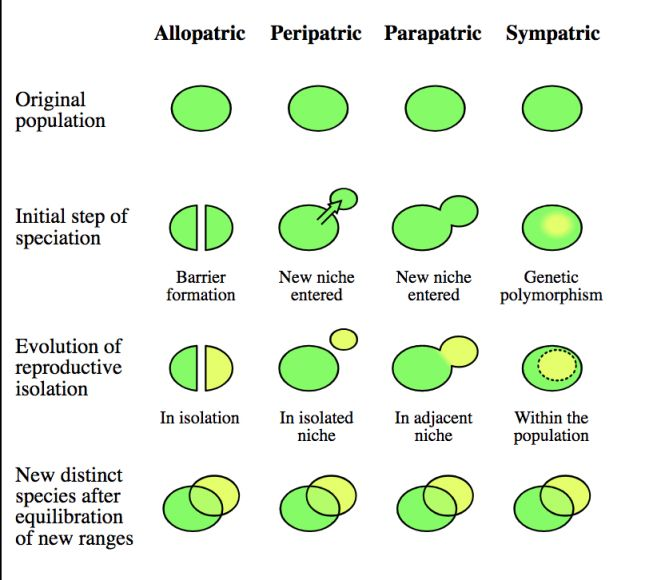
- Аллопатрия (alloparty) — это означает «в другом месте», и описывает популяцию или вид, который физически изолирован от других подобных групп внешним барьером для расселения. С точки зрения биогеографии, аллопатрические виды и популяции – это те, которые не имеют перекрывающихся географических ареалов.
- Генетическая несовместимость (genetic incompatibility) – процесс, при котором в результате спаривания получается потомство, нежизнеспособное, склонное к заболеваниям или генетически дефектное в каком-либо отношении.

Новые адаптивные зоны
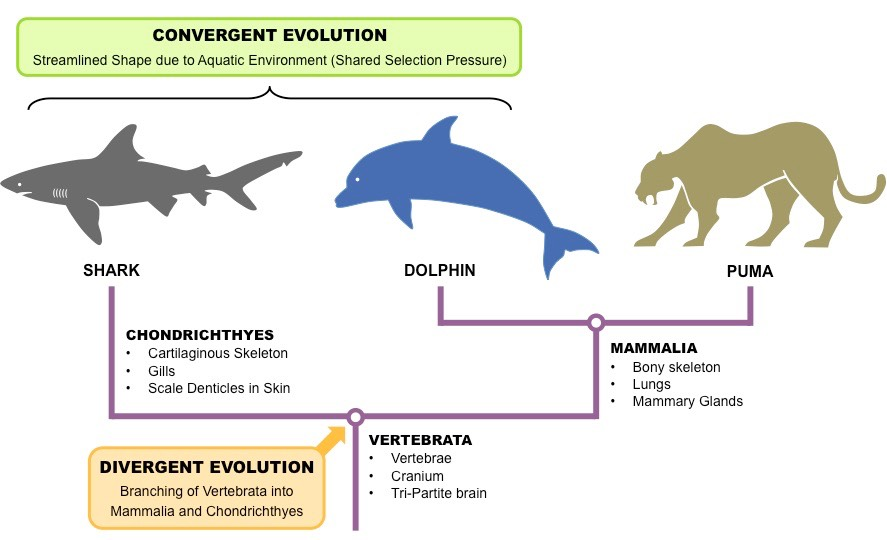
На этом уровне изменений открываются совершенно новые возможности и типы живых существ. Например, колонизация суши амфибиями ~370 Ма (Ma–million years ago) или независимое появление гомойотермности (терморегуляция, поддерживающая стабильную внутреннюю температуру тела независимо от внешних условий) у млекопитающих и птиц ~250 Ма. Есть не столь эпохальные, но тем не менее масштабные инновации, как, например, эхолокация у китообразных и летучих мышей, или жвачный желудок у парнокопытных животных. Это конвергентная эволюция – независимое развитие сходных признаков или особенностей (например, строения тела или поведения) у неродственных или отдаленно родственных видов или родов, которые обычно занимают сходную среду или экологические ниши.
Большие переходы (major transitions)
Концепция больших переходов сформулирована в книге Джона Мэйнарда Смита и Эорса Шатмари «The Major Transitions in Evolution«, в которой они определяют общие свойства для их определения:
- Более мелкие сущности часто объединяются вместе, образуя более крупные. Например, хромосомы, эукариоты, многоклеточные колонии.
- Более мелкие сущности часто становятся дифференцированными как часть более крупной сущности. Например, ДНК и белок, органеллы, анизогамия, ткани, касты.
- Более мелкие образования часто не способны воспроизводиться в отсутствие более крупного образования. Например, ДНК, хромосомы, органеллы, ткани, касты.
- Меньшие сущности иногда могут нарушить развитие более крупной сущности, например, мейотический драйв (эгоистичные неменделевские гены), партеногенез, рак, государственный переворот.
- Возникли новые способы передачи информации, например, ДНК-белок, клеточная наследственность, эпигенез, универсальная грамматика.
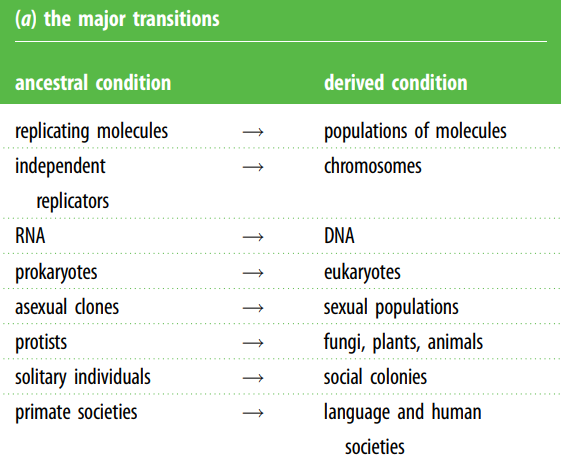
Ключевым элементом больших переходов является повышение сложности и изменение систем передачи информации. Эукариотическая клетка сложнее прокариотической, половое размножение сложнее бесполого и т.д. Крупные переходы — это такие переходы, при которых происходит изменение уровня организации, последствия которого способны изменить правила жизни. При крупных переходах субъекты, которые ранее воспроизводились независимо, впоследствии воспроизводятся как часть более крупной единицы, что может повлиять на единицы и уровни отбора. Такое изменение может привести к специализации (и, следовательно, к разнообразию функций в организме) и к изменению способа передачи информации между поколениями.
А что же люди?
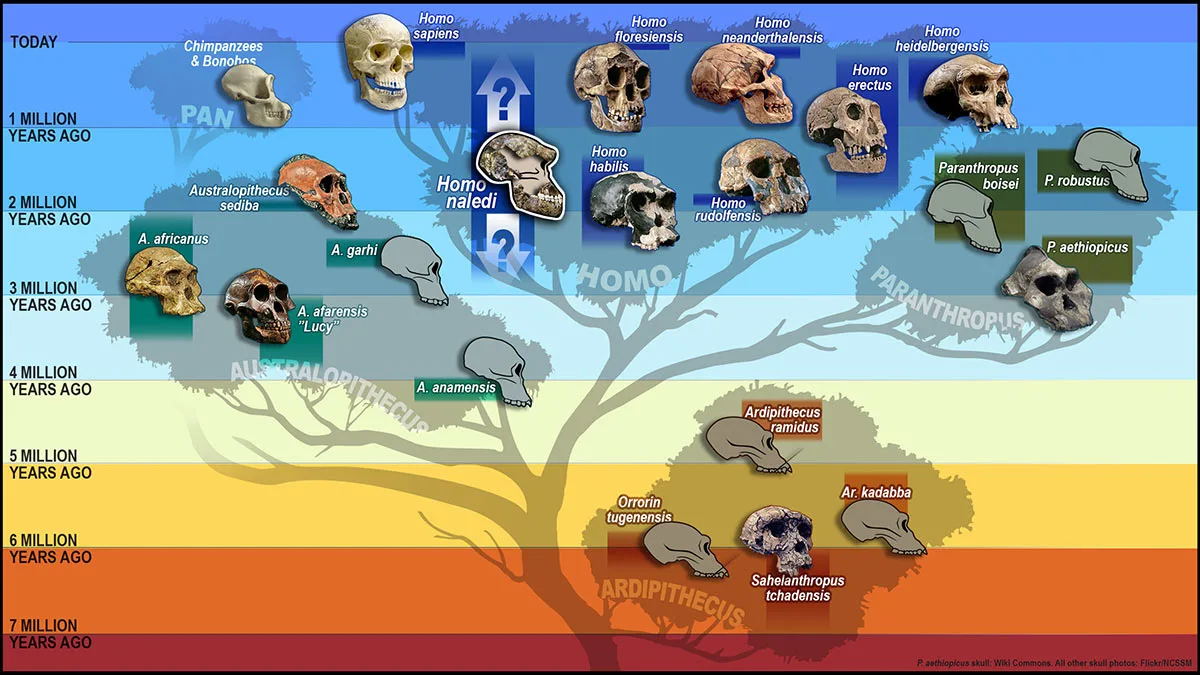
Совершила ли эволюция большой переход, результатом которого стали мы? Возможно. Шатмари утверждает: «Биология дает простор для технологической и коллективной (communal) культурной эволюции. Благодаря социальной заботе (включая медицину) и сельскому хозяйству биология человека постепенно дедарвинизировалась. Именно культура является тем местом, где происходит основное действие». По сути, культура заменяет биологию в качестве главной области изменения и отбора. Посмотрим на изменения, которые происходили начиная с самого низкого эволюционного уровня.
Базовая эволюция человека

Имеется достаточно много физических доказательств градуальной эволюции человека. Нетрудно догадаться, в каком виде, если учесть, что они сохранились на протяжении миллионов и сотен тысяч лет – это кости и зубы. Изменения в размерах и формах тела, мозга и зубов, могут многое рассказать. Но есть, как обычно, сложность, даже две. Во-первых, изменения в этих параметрах не всегда однонаправленные (подтверждение тому Homo floriensis, которые уменьшились в размерах). А во-вторых, палеоантологическая летопись окаменелостей (paleoantological fossil record) прерывиста и неполна – останков не так много в общем, и в рамках каждого отдельного вида в частности. Тем не менее, увеличение нейрокраниума (мозговой части черепа) с 400см3 до 1400см3, как и динамика изменения зубов, размеров тела, костей лицевой части черепа, четко прослеживаются. Молекулярные подходы, в свою очередь, оперируя частотой мутаций, свидетельствуют в пользу кумулятивной природы мелкомасштабных изменений.
Образование новых таксонов человека
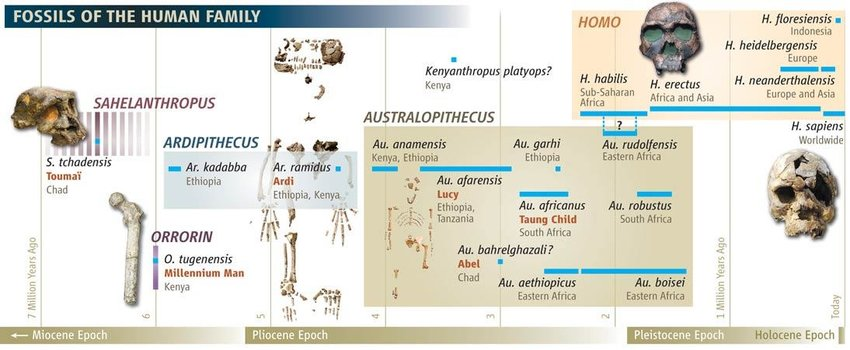
Чтобы говорить о двух новых видах там, где раньше был один, нужно определение вида. С биологической концепцией, которая подразумевает наличие репродуктивных барьеров между двумя генетическими пулами, будут сложности – мы не можем с точностью сказать, как экология разделяла популяции в далёком прошлом. На этот случай есть альтернативная концепция эволюционного вида: линии, для которой есть доказательства независимой эволюционной траектории.
Есть три параметра, которые помогут прочесть палеонтологическую летопись. Это дата первого появления окаменелостей (first appearance date, FAD), дата последнего появления окаменелостей (last appearance date, LAD) и количество таксонов на момент времени, или разнообразие (Divercity). Их можно рассматривать, как условные показатели видообразования, вымирания и видового разнообразия.
Пики на графиках отражающих эти три параметра рассказывают свою историю. Мы видим несколько пиков видообразования, самый большой из которых произошёл 2–2.5 Ma, и пики вымирания, самый большой 2,0-1,5 Ma, с меньшими пиками 3,0–3,5 Ma и 0–0,5 Ma. Главная мысль здесь в том, что макроэволюционный паттерн отражает скорее кумулятивные изменения на протяжении длительного срока, нежели короткие периоды интенсивных изменений. Это, однозначно, не просто градуальное анагенетическое изменение.
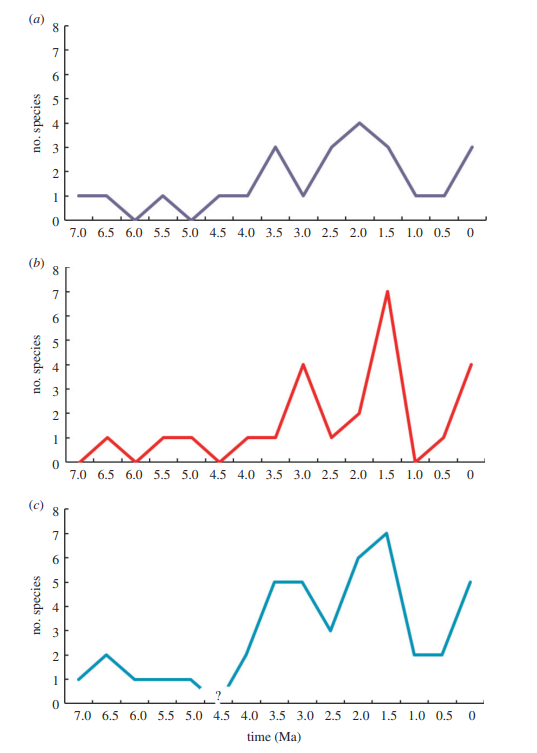
Пик вымираний 2,0–1,5 Ma идёт после пика видообразования – это влияние эволюции и распространения рода Homo на другие формы гоминин. А график разнообразия говорит об адаптивной радиации гоминин в это время, пусть и кратковременной. Было много разных, но недолго.
Обратите внимание на последний пик 0,5 Ma когда есть и появления, и вымирание и разнообразие. Это период, когда люди эволюционировали вместе с другими представителями рода Homo. Евразийские Неандертальцы и Денисовцы сосуществовали с африканскими Сапиенсами одновременно, вплоть до 30 ka (kilo annum, тысяч лет назад), когда остались только мы.
Новая адаптивная зона человека
Третий уровень перехода подразумевает новую адаптивную зону или значительные адаптивные изменения. Одним различием в размерах тела или зубов тут не обойтись. С момента отделения от общего предка с шимпанзе около 5–6 Ma, прошли три этапа изменений, сравнимые по масштабности с освоением суши.
Первый, плиоценовый переход ~5-4 Ma,, по-видимому (насколько это можно показать), был связан с паттернами локомоции (то есть целенаправленного передвижения) и варьирующего поведения (ranging behavior), предполагая новую среду обитания и экологическую нишу. Вероятно, они изменялись по мере того, как в окружающей среде стали преобладать леса и луга. Неизбежно должны были произойти изменения в диете, поведении и социоэкологии, чтобы адаптироваться к другому окружению.
Второй, плио-плейстоценовый переход, который длился долгих два миллиона лет, сложнее и гораздо лучше документирован. В этот период происходят изменения в диете, способах добычи ресурсов (появляется технология), изменяется жизненная стратегия (life-history strategy). Самыми ранними элементами этого перехода является появление каменных орудий в Ломекви, датируемое 3,3 Ма; другие элементы включают первые свидетельства обработки животных с помощью орудий (3,4 Ма); появление рода Homo, или, точнее, фенотипов, ассоциируемых с родом человеческой линии, а именно: более крупный мозг, уменьшенные постканинные зубы (моляры и премоляры), менее прогнатическое (выступающее) лицо и развитие характерных прерывистых супраорбитальных (надбровных) дуг.
Примерно 1,8 Ма происходит значительное увеличение числа стоянок и размера коллекций инструментов, что свидетельствует о переходе к более привычной модели использования орудий. Примерно в это же время увеличивается количество свидетельств разделывания животных, возможно, в результате охоты. Также, по-видимому, в этот период происходили первые расселения в северной Африке и Евразии. Главным моментом, который следует подчеркнуть для этого сложного перехода, является то, что это не единичное событие в сжатый период времени, а растянутое более чем на миллион лет множество мелких микроэволюционных сдвигов.
На третий переход позднего четвертичного периода (Latest Quaternary, это Антропоген, текущий период кайнозойской эры) приходятся серьезные поведенческие, когнитивные и культурные изменения. В течение всего периода наблюдается значительное увеличение размера мозга, изменения в морфологии черепа и общей прочности, но по сравнению с прогрессом в более ранние переходные периоды, они относительно незначительны. Ключевые элементы этой фазы: ускорение темпов изменений и повышение сложности технологий, возникновение региональных образований и идентичностей, увеличение плотности населения, интенсификация культурных процессов, символическое мышление и репрезентация. А скорость этих изменений просто головокружительна в масштабах земных процессов – этот период занял менее 0,5 Ма, что намного меньше нескольких миллионов лет двух других переходов.
Обеспечивают ли все описанные эволюционные изменения вместе взятые большой переход, в том смысле, какой в него вкладывали Мэйнард Смит и Шатмари? Критерием здесь является появление более крупных сущностей репликации, новых путей передачи информации, разделение ролей и потеря независимости репликации, которая приводит к эволюционной хрупкости.

Глобальный эволюционный переход человека может быть вызван технологической зависимостью, языком, кумулятивной культурой, высоким уровнем репродуктивного сотрудничества и кооперацией между неродственными особями. Они все взаимосвязаны и не представляется возможным вычленить какой-то из этих феноменов в качестве ключевого. Как и на более низких уровнях эволюционных изменений, видно, что эволюция человека это не единичное событие, и не серия резких “революционных” скачков, а мозаичный процесс комбинирования и аккумулирования, где мозаичность заключается в том, что разные черты появляются и эволюционируют независимо и в разное время.
