
Оригинал статьи Evidence for a Large-Scale Brain System Supporting Allostasis and Interoception in Humans
Мозг содержит внутренние системы обработки экстероцептивных сенсорных сигналов из окружающей среды, таких как зрение, слух и проприорецепция/осязание. (Sepulcre, J.,et al., 2012). Накопилось достаточно свидетельств того, что эти системы функционируют согласно принципу прогностической обработки (predictive coding), в котором ощущения предвосхищаются (anticipated) и затем корректируются сенсорным вводом (sensory input) из среды (Rao, R. P. & Ballard, D. H., 1999; Chennu, S. et al., 2013; Adams, R. A., Shipp, S. & Friston, K. J., 2013; Kusumoto-Yoshida, I., Liu, H., et al., 2015).
Мозг, как генеративная система, моделирует мир скорее предсказывая, чем реагируя на сенсорный ввод. Предсказания направляют действия и восприятие постоянно конструируя возможные репрезентации непосредственного будущего основываясь на своих предыдущих вероятностных оценках (prior probabilities) в зависимости от текущего контекста (Clark, A., 2013; Friston, K., 2010). Ряд исследовательских коллективов изучает гипотезу о том, что восходящий поток сенсорных данных (ascending sensory inputs) от органов и систем внутренней среды организма таким же образом предвосхищается и репрезентируется (автономная висцеральная и васкулярная функции, нейроэндокринные флуктуации и нейроиммунная функция) (Pezzulo, G., Rigoli, F. & Friston, 2015; Barrett, L. F. & Simmons, W. K., 2015; Seth, A. K., 2013; Seth, A. K., Suzuki, K. & Critchley, H. D., 2012; Gu, X. & FitzGerald, T. H., 2014; Allen, M. & Friston, K. J., 2016). Эти ощущения называются интероцепцией (Craig, A. D., 2002).
Технические исследования нейронных структур (Sterling, P. & Laughlin, S., 2015) наряду с физиологическими доказательствами (Sterling, P., 2012), указывают на то, что мозг непрерывно предвосхищает энергетические потребности тела и готовится к их удовлетворению еще до того, как они возникнут. Этот процесс называется аллостазом.
Аллостаз, это не условие или состояние — это процесс, посредством которого мозг эффективно поддерживает энергетическую регуляцию тела. Аллостаз определяется в терминах прогнозирования (prediction), и современные теории предполагают, что предсказание интероцептивных сигналов необходимо для успешного аллостаза (Barrett, L. F., 2016; Barrett, L. F., 2017). Вдобавок к восходящим путям и отделам мозга важным для интероцепции, актуальные теоретические дискуссии предполагают существование распределенной аллостатически-интероцептивной системы в мозге (аналогичной экстероцептивным системам).
В экспериментах на макаках получены анатомические субстраты для гипотезы о том, что в мозге есть единая внутренняя система для аллостаза и интероцепции (Morecraft, R. J. et al., 2012). В данной работе, с использованием анализа функциональных связей в состоянии покоя с помощью фМРТ, получены доказательства наличия этой предполагаемой системы у человека.
Анализ эмпирических данных демонстрирует, что аллостатически-интероцептивная система является общедоменной (domain-general) и поддерживает широкий диапазон психологических функций включая интероцепцию, эмоции, память, вознаграждение и когнитивный контроль (Barrett, L. F. & Satpute, 2013). Таким образом, чем бы эта система ни была занята — запоминанием, направлением внимания и тому подобным — она также прогностически регулирует физиологические системы тела поддерживая аллостаз для реализации этих функций (Barrett, L. F., 2017; Barbas, H. & Rempel-Clower, N., 1997).
Анатомическое обоснование существования аллостатически-интероцептивной системы
Более трёх десятилетий исследований нервных путей мозга макак (tract-tracing studies) продемонстрировали анатомические субстраты предполагаемого потока предсказаний и сигналов ошибок предсказаний (prediction error signals). В работе Barbas, H. & Rempel-Clower, N., 1997 показан поток информации между ламинарными градиентами специфичных областей в соответствии со структурной моделью кортико-кортикальных связей, подробнее у Barbas, H., 2015. Вдобавок, эта структурная модель органично встроена в систему прогностической обработки (Barrett, L. F. & Simmons, W. K., 2015; Chanes, L. & Barrett, L. F., 2016).
В отличие от других моделей потока информации в коре мозга, структурная модель успешно предсказывает поток информации в лобной, височной и затылочной областях коры (Rempel-Clower, N. L. & Barbas, H., 2000; Medalla, M. & Barbas, H., 2014; Medalla, M. & Barbas, H., 2006). Прогностические сигналы идут от областей с низкой ламинарностью (агранулярные области) к слоям с высокой ламинарностью (гранулярные области), в то время как сигналы прогностических ошибок идут в обратном направлении.
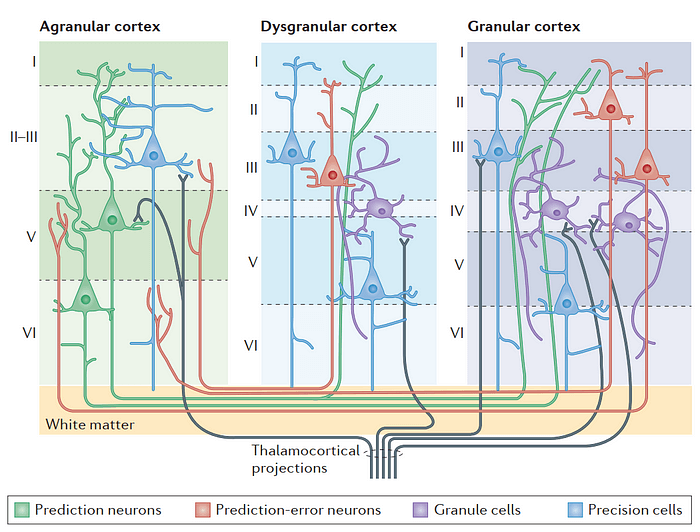
Активный вывод (active inference) в коре
Кортикальные колонки определяются количеством слоев (laminae), где каждый слой характеризуется отдельными типами клеток и паттернами интра- и интеркортикальных связей. Гранулярная кора (справа) характеризуется шестью дифференцированными слоями (слои I–VI), где IV слой содержит гранулярные клетки — возбуждающие звездчатые шипиковые нейроны (лиловым) — которые усиливают и распространяют входящий таламокортикальный сигнал по колонке. Гранулярная кора также содержит много шипиковых пирамидальных нейронов в своих инфра- и супрагранулярных слоях. У пирамидальных нейронов есть: треугольная сома, от которой отходят базальные дендриты; восходящий апикальный дендрит, зачастую c большим пучком дендритов в I слое; и нисходящий аксон, который выходит из колонки (иногда с несколькими коллатералями). Для сравнения, агранулярная кора (слева) не имеет полностью выраженного IV слоя и обладает слабодифференцированной границей между II и III слоями. Эти верхние слои содержат сравнительно меньшее количество пирамидальных нейронов чем гранулярная кора. Однако, агранулярная кора содержит относительно бОльшее количество в слоях V и VI чем в своих верхних слоях. Не смотря на отсутствие выраженного IV слоя с гранулярными клетками, агранулярная кора все равно имеет таламические проекции; однако, сенсорная информация, которая приходит в агранулярную кору не так усилена и хуже распределена по колонке чем в гранулярной коре. Дисгранулярная кора находится в промежуточных зонах между гранулярными и агранулярными областями и содержит небольшой но выраженный IV слой и характерные (хоть и рудиментарные) слои II и III. Эта иллюстрация (Рис. 1) не претендует на исчерпывающую демонстративность, но подчеркивает ламинарные и клеточные характеристики важные для понимания Embodied Predictive Interoception Coding (EPIC) модели и ее предсказаний. В соответствии с ней, прогностические нейроны (изображенные как зеленые пирамидальные нейроны) в глубоких слоях агранулярной коры направляют активный вывод путем отправки сенсорных предсказаний через проекции (зеленые линии) в супрагранулярные слои дисгранулярных и гранулярных сенсорных участков. Нейроны ошибок предсказаний (error prediction neurons) (изображены красными пирамидальными нейронами) в супрагранулярных слоях гранулярной коры вычисляют разницу между предсказанным и полученным сигналами, и посылают сигналы ошибки предсказания через проекции (красные линии) обратно в глубокие слои агранулярных кортикальных слоев. Прецизионные клетки (precision cells) (изображены синими пирамидальными нейронами) “подстраивают чувствительность” к предсказаниям и ошибкам предсказания, придавая таким образом этим сигналам сниженный (или, в некоторых случаях, повышенный) “вес” в зависимости от относительной уверенности в нисходящих предсказаниях или надежности входящих сенсорных сигналов.
В разработанной теории интероцепции, EPIC (Embodied Predictive Interoception Coding) модели, подход “активного вывода” Карла Фристона (active inference) к прогностической обработке интегрирован со структурной моделью Хелен Барбас, для предположения, что менее дифференцированные агранулярные и дисгранулярные висцеромоторные участки в поясной коре и передней островковой коре инициируют висцеромоторные предсказания через свой каскад связей в гипоталамус, периакведуктальное серое вещество и другие ядра ствола мозга контролирующие внутреннюю среду (Weston, C. S., 2012; Vogt, B. A., 2005) (красные пути на рис. 2); одновременно с этим, поясная кора и передняя островковая кора посылают ожидаемые сенсорные последствия этих висцеромоторных действий (а именно, интероцептивных предсказаний) в более гранулярную первичную интероцептивную кору в дорсальной части средней и задней островковой коры (dmIns/dpIns, синие пути, сплошная линия, на рис. 2) (Nieuwenhuys, R., 2012).
Следуя этой логике, был обнаружен набор ключевых кортикальных регионов c висцеромоторными связями, которые должны формировать основу общедоменной системы интероцепции и аллостаза. Как предполагается EPIC моделью, бОльшая часть ключевых висцеромоторных отделов в предполагаемой интероцептивной системе имеют, фактически, моносинаптические, двунаправленные связи с первичной интероцептивной корой, поддерживая гипотезу о том, что они напрямую обмениваются интероцептивными предсказаниями и сигналами ошибки предсказаний. Это висцеромоторные кортикальные регионы моносинаптически проецируются в субкортикальные и стволовые отделы контролирующие внутреннюю среду (то есть, автономная нервная система, иммунная и нейроэндокринная системы), такие как гипоталамус, периакведуктальное серое вещество, парабрахиальное ядро, вентральный стриатум и ядро одиночного пути (красные пути на рис 2.).
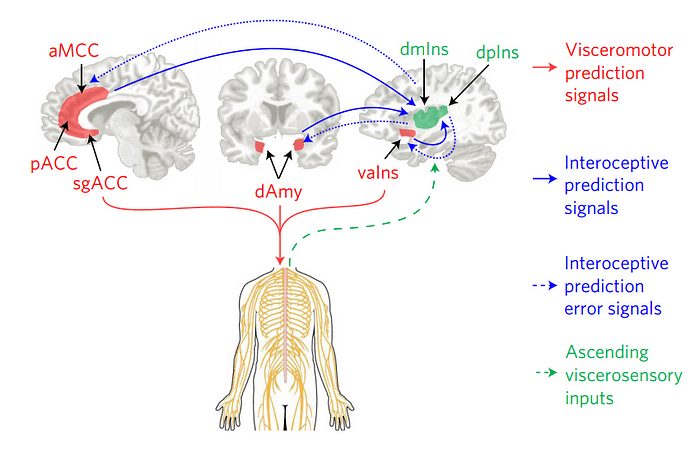
Отделы с менее развитой ламинарной организацией отправляют ожидаемые сенсорные последствия висцеромоторных изменений (интероцептивные прогностические сигналы) в первичную интероцептивную кору с более развитой ламинарной организацией (сплошная синяя линия). Входящие сенсорные сигналы из внутренней среды проводятся по блуждающему нерву и волокнам небольшого диаметра C и Aδ (прерывистая зеленая линия) в первичную интероцептивную кору в дорсальном секторе средней и задней островковой коры. Текущие интероцептивные предсказания могут быть актуализированы передачей ошибок предсказания в висцеромоторные регионы (прерывистые синие линии).
Функционирование аллостатически-интероцептивной системы
Интегрированная аллостатически-интероцептивная система мозга является сложной кортикальной и субкортикальной системой состоящей из взаимосвязанных внутренних сетей. Эта единая система не только обеспечивает аллостаз, но и целый ряд психологических феноменов (эмоции, память, принятие решений, боль), что может быть объяснено их зависимостью от аллостаза. Фактически, такова было логика Пейпеца, предполагавшего, что функционально лимбическая система “предназначена” для эмоций. Данная работа идет еще дальше. Отделы контролирующие внутреннюю физиологию тела являются также компонентами нейронных сетей обеспечивающих социальную принадлежность, боль, суждения, эмпатию, вознаграждение, аддикцию, память, стресс, тягу (craving) и принятие решений (рис 3). Всё больше работ по функциональной нейровизуализации обнаруживают общедоменность (domain-general) дефолт-системы (default-mode network) и салиентной системы (salience network) (Barrett, L. F. & Satpute, A. B., 2013; Anderson, M. L., 2014; Yeo, B. T. et al., 2015).
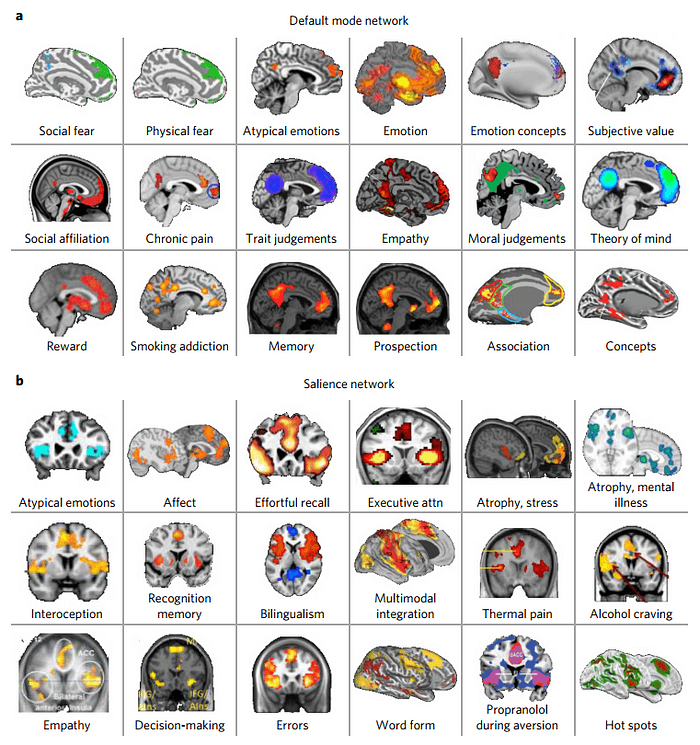
Согласно гипотезе EPIC модели висцеромоторный контроль и интероцептивные входящие сигналы интегрированы в единую систему, в отличие от традиционных взглядов на кортикальные отделы посылающие висцеромоторные сигналы и отделы получающие интероцептивные сигналы как две отдельные системы, по аналогии с кортикоспинальной скелетодвигательной эфферентной системой и первичной соматосенсорной афферентной системой.
Возможно, самое важное то, что аллостатически-интероцептивная система играет свою роль в ряде психологических феноменов. Можно предположить, что аллостаз и интероцепция являются фундаментальными свойствами нервной системы. Анатомические, физиологические и вычислительные (signal-processing) доказательства предполагают, что мозг не эволюционировал для рациональности, счастья или точной перцепции; скорее,
все мозги выполняют одну главную функцию: эффективно обеспечивать и распределять ресурсы для физиологических систем в организме животного (его внутренней среде) в целях роста, выживания, развития и репродукции
(Sterling, P. & Laughlin, S., 2015).
То есть, мозг эволюционировал для регулировки аллостаза (Sterling, P., 2012). Все психологические функции работающие на рост, выживание, развитие и репродукцию (такие как ощущение эмоций, воспоминание, направление внимания или принятие решений) требуют эффективного регулирования метаболических и других биологических ресурсов.
Комплекс салиентной и дефолт-системы в EPIC модели служит мощной основой для интеграции информации во всём мозге (van den Heuvel, M. P., Kahn, R. S., Goni, J. & Sporns, 2012). Диффузионная тензорная визуализация показала, что эти системы включают самое большое количество хабов относящихся к “клубу богачей” мозга, содержащих наибольшее количество связей в коре (van den Heuvel, M. P. & Sporns, O., 2011; van den Heuvel, M. P. & Sporns, O., 2013), некоторые из которых являются коннекторными хабами в аллостатически-интероцептивной системе (см. Рис 4). Все остальные сенсорные и моторные сети взаимодействуют с дефолт- и салиентной системами, и возможно, между собой, через эти хабы (Sepulcre, J., et al., 2012; van den Heuvel, M. P. & Sporns, O., 2013).
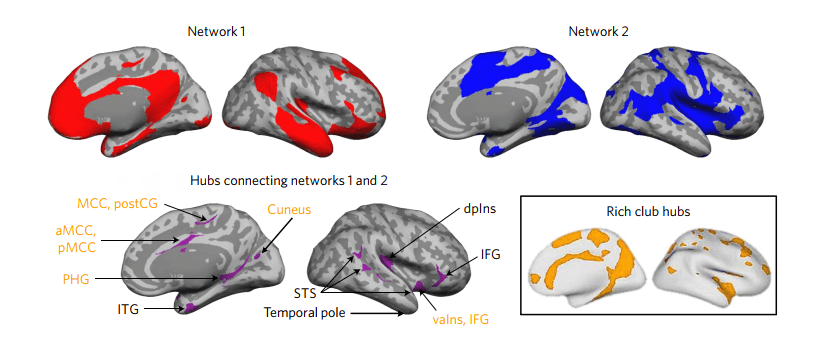
Сети показаны красным и синим, хабы изображены лиловым; хабы относящиеся к “клубу богачей” изображены желтым.
Важно то, что полученные данные позволяют внести ясность в психологические концепты, которые часто путали в психологической и нейронаучной литературе: аффект и эмоции. Чем бы ваш мозг ни был занят — думал, чувствовал, воспринимал, “двигался” — он также регулирует вашу автономную нервную систему, иммунную систему, эндокринную систему, и в то же время непрерывно репрезентирует интероцептивные последствия этих физических изменений. Интероцептивные ощущения обычно воспринимаются как ощущения аффекта низкого разрешения (low-dimensional) (Barrett, L. F., 2017; Barrett, L. F. & Bliss-Moreau, E., 2009).
Одним из наиболее важных аспектов этой работы является выход за рамки традиционного общедоменного или “модульного” взгляда на структурно-функциональную организацию мозга, который предполагает значительную степень специфичности отдельных систем (Fodor, J. A. The Modularity of Mind, 1983).
Растущий объем доказательств требует отхода от этого традиционного модульного подхода (Li, D., Christ, S. E. & Cowan, N., 2014; Fuster, J. M., 2000) в пользу моделей признающих, что нейронные популяции являются общедоменными (domain-general). Идея общедоменности даже применима к первичным сенсорным сетям. Это подтверждается тем, что мультисенсорная обработка происходит в отделах мозга традиционно считавшихся унимодальными (например, слуховая кора отвечающая на визуальную стимуляцию, Kayser, C., Petkov, C. I., Augath, M. & Logothetis, N. K., 2007; Liang, M., Mouraux, A., Hu, L. & Iannetti, G. D.).
И тут еще один очень любопытный момент, в статье есть следующий пассаж “Отсутствие специфичности в отношениях структуры и функций мозга не является мерой ошибки или биологической дисфункции, но полезным свойством отражающим основные принципы биологической дегенерации оставившими след в геноме, иммунной системе и любой другой биологической системе сформированной естественным отбором (Edelman, G. M. & Gally, J. A. Degeneracy and complexity in biological Systems, 2001). Кто дочитал досюда и захочет глубже изучить вопрос, есть интересная ссылка (Degeneracy: a link between evolvability, robustness and complexity in biological systems, James M Whitacre, 2010).
