Приблизительно 8% человеческого генома представляет себя из себя ретровирусы на разной стадии “фоссилизации”. Когда-то они инфицировали наш геном, чтобы на чужом горбу въехать в далекое светлое будущее, да так и остались. Обычно большинство подобных генетических вторжений либо (i) настолько разрушительны и фатальны, что просто не передаются дальше, либо (ii) не оказывают особого влияния и со временем “стачиваются” в процессе накопления мутаций.
Хотя большая часть ретровирусных вкраплений побита мутационной дегенерацией как молью, некоторые ретровирусные гены удивительным образом сохранились вопреки мутантной инактивации. Это редкие случаи узурпирования организмами ретровирусных генов в своих целях. Особенно интригует “одомашнивание” envelope (env) генов. Самые известные из них, синцитины, были кооптированы геномом хозяина чтобы выступать посредником в обмене питательными веществами между матерью и эмбрионом у плацентарных млекопитающих. Самый старый из них, ген Syncytin-Car1 gene, был “одомашнен” около 60 миллионов лет назад, до отделения эволюционной ветви Хищников (Carnivora).
В контексте ретровирусных генов, пресловутые env гены кодируют протеиновый прекурсор, который “вонзается” в поверхностные (surface, SU) и трансмембранные (transmembrane, TM) белки позволяющие вирионам прикрепляться к таргетным клеткам и проникать в них мембранным фьюжном (слиянием). SU белки служат для прикрепления вириона к клетке-хозяину путем взаимодействия с рецепторами на поверхности клеточной мембраны, а ТМ белки служат для заякоривания всего гликопротеинового комплекса на поверхности вириона и обеспечения слияния вириона с мембраной клетки-хоста (хозяина). Многие env гены кодируют также пептидный мотив в ТМ домене, который имеет иммуносупрессорные свойства и подавляет производство цитокинов и клеточный иммунитет. Примечательно то, что эти две функции — клеточный фьюжн и уклонение от иммунной системы — ключевые для выживания плода у многих млекопитающих. Предположительно, envelope-ассоциированные иммуносупрессивные свойства могут защищать плод от иммунной атаки материнской иммунной системы.
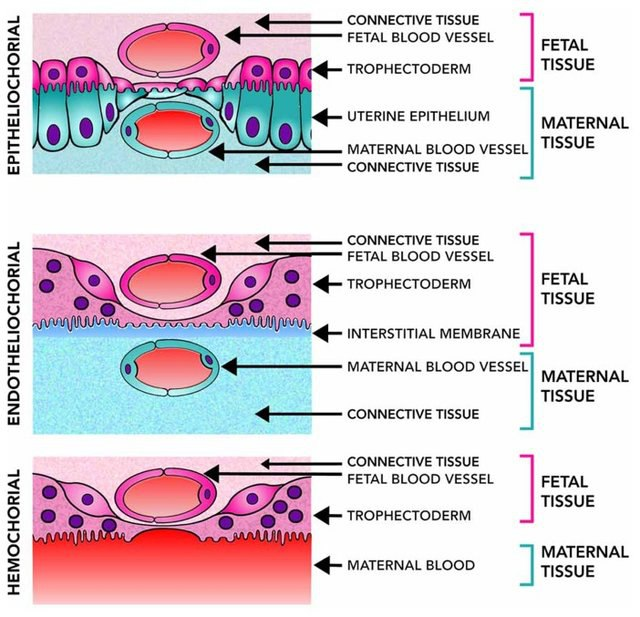
У млекопитающих существует три плацентарных формы. При гемохориальном типе плаценты — у высших приматов, зайцеобразных и грызунов — хорион омывается материнской кровью, При эндотелиохориальной форме, например, у хищников, хорион контактирует с эндотелиальной оболочкой материнских сосудов. А при эпителиохориальной плацентации — с эпителием матки. В этом третьем случае не образуется синцитиотрофобласт, и отличие от других двух типов, гены синцитины могут вообще не задействоваться. Филогенетическая реконструкция свидетельствует о том, что наиболее инвазивная (т.е. гемохориальная) форма плацентации, при которой образуется синцитиотрофобласт, является “праотеческой” для всех плацентарных млекопитающих.
Таким образом, предполагается, что разнообразные морфологические инновации в плацентации у млекопитающих (даже среди гемохориальных форм) могут быть обусловлены приобретением разных генов синцитинов, с разными рецепторами и разными фьюзогенными свойствами (способностью к слиянию).
В то время, как оптимальная эволюционная стратегия для генов матери — распределение ресурсов равномерно среди всего потомства, чтобы максимизировать чистую эволюционную выгоду, каждое отдельное потомство (а именно его отцовские гены) пытается отжать у матери больше ресурсов. В контексте этого генетического конфликта за материнские ресурсы кооптация каждого нового синцитина может обеспечивать плоду дополнительные возможности урвать побольше материнского питания. Возможно, что колебания уровня напряженности этого конфликта стали драйвером развития разных типов плацентации у млекопитающих, сопровождаясь приобретением или потерей синцитиновых генов.
Retroviruses push the envelope for mammalian placentation
